

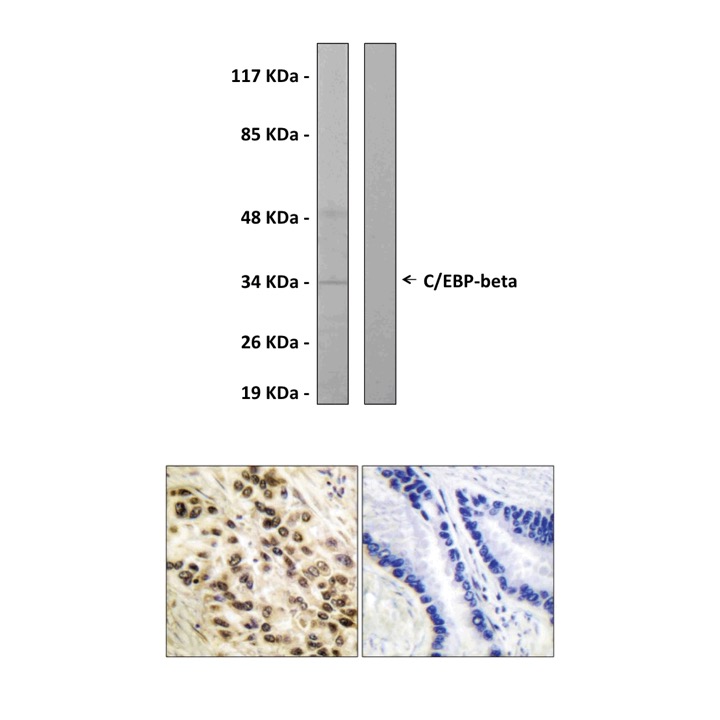
Anti-C/EBP-β: Rabbit C/EBP-beta Antibody |
 |
BACKGROUND C/EBPbeta is a member of the bZIP family of transcription factors that binds to DNA as homodimers or as heterodimers with other C/EBPs. Three isoforms of C/EBPbeta (38kD, 34kD, and 20kD in mice) are produced to various extents in different cell types and at specific stages of differentiation through alternative use of three translation initiation sites within a single mRNA. The most ubiquitous isoform (34kD), usually referred to as LAP (liver-enriched transcriptional activator protein), is composed of a C-terminal region containing the basic DNA-binding and leucine zipper dimerization domains and a N-terminal region, which contains the transactivation domains. A larger isoform (38kD), expressed in a differentiation-associated manner in some cell types including preadipocytes, is identical to LAP except that it contains an additional transactivation domain at the N terminus. Both LAP and the 38kD protein are potent transactivators of gene expression, whereas the 20kD isoform, LIP (liver-enriched transcriptional inhibitory protein), represses transcription in a dominant negative manner.1
The transcriptional activity of C/EBPbeta is regulated by several mechanisms, including association with other proteins as well as posttranslational modification.2 It has also been suggested that C/EBPbeta normally exists in a repressed state and is activated by phosphorylation of a repressor domain located between the N-terminal transactivation domains and the C-terminal bZIP region. This domain contains several serines and threonines, all of which are phosphorylated to some extent by a constitutive process. The site encompassing threonine 188 (SPPGT188PSP) in murine LAP is a consensus site for both glycogen synthase kinase 3 (GSK3) and extracellular signal-regulated kinase (ERK), which is dephosphorylated in response to growth hormone stimulation of preadipocytes. Modification of this site by mutation of the threonine to alanine inhibits the C/EBPbeta-associated transcription of a c-fos promoter reporter in response to growth hormone signaling.3
Recent Publication
Kim, K.S., H.M. Choi, H.I. Ji, C. Kim, J.Y. Kim, R. Song, S.M. Kim, Y.A. Lee, S.H. Lee, H.I. Yang, M.C. Yoo, and S.J. Hong. 2013b. Effect of taurine chloramine on differentiation of human preadipocytes into adipocytes. In Advances in experimental medicine and biology. Vol. 775. 247-257.
REFERENCES
1. Descombes, P. et al: Cell 67:569-79, 1991
2. Park, B.H. et al: Mol. Cell. Biol. 24:8671-80, 2004
3. Piwien-Pilipuk, G. et al: J. Biol. Chem. 277:44557-65, 2002
2. Park, B.H. et al: Mol. Cell. Biol. 24:8671-80, 2004
3. Piwien-Pilipuk, G. et al: J. Biol. Chem. 277:44557-65, 2002
Products are for research use only. They are not intended for human, animal, or diagnostic applications.
Параметры
Cat.No.: | CG1055 |
Antigen: | Synthesized peptide derived from human C/EBP-β |
Isotype: | Rabbit IgG |
Species & predicted species cross- reactivity ( ): | Human, Mouse, Rat |
Applications & Suggested starting dilutions:* | WB 1:500-1:1000 IP n/d IHC 1:50-1:100 ICC n/d FACS n/d |
Predicted Molecular Weight of protein: | 36 KDa |
Specificity/Sensitivity: | Detects endogenous C/EBP-β proteins without cross-reactivity with other family members. |
Storage: | Store at -20°C, 4°C for frequent use. Avoid repeated freeze-thaw cycles. |
*Optimal working dilutions must be determined by end user.
Документы

Информация представлена исключительно в ознакомительных целях и ни при каких условиях не является публичной офертой



