BACKGROUND
Protein Kinase C (PKC) is a family of serine- and threonine-specific protein kinases that can be activated by calcium and the second messenger diacylglycerol. PKC family members phosphorylate a wide variety of protein targets and are known to be involved in diverse cellular signaling pathways. They also serve as major receptors for phorbol esters, a class of tumor promoters. Each member of the PKC family has a specific expression profile and is believed to play a distinct role in cells. The PKC family can be broadly divided into three groups that differ in their cofactor requirements. These are known as conventional (c)PKC isoforms (alpha, betaI, betaII and gamma), that require Ca2+ and diacylglycerol (DAG) to become activated; novel (n)PKC isoforms (delta, epsilon, zeta, theta and mu) that require only DAG; and the atypical (a)PKC isoforms, namely zeta, iota and lambda (the mouse homologue of human PKCiota), that require neither Ca2+ nor DAG.1 All PKC family members possess a phosphatidylserine (PS) binding domain for membrane interaction. All PKCs isoforms consist of a highly conserved catalytic domain connected to a regulatory domain via a hinge region. The general structure of a PKC molecule consists of a catalytic and a regulatory domain found at the C- and N-terminus respectively. Both domain structures are composed of a number of conserved regions (C1–C4) interspersed with regions of lower homology, so-called variable domains, V0–V5. Under baseline conditions, PKC is rendered largely inactive by intramolecular interaction between ‘pseudosusbstrate’ and catalytic sites, which causes PKC autoinhibition. PKC activation weakens conformational interactions between the pseudosubstrate and the catalytic sites, and makes the pseudosusbstrate sequences less accessible to the PKC catalytic moiety.2 Calcium-dependent PKC isoforms have been shown to be stimulated by diacylglycerol, in the presence of calcium ions and phospholipids. Diacylglycerol, containing unsaturated fatty acids, was identified as the most active constituent, capable of enhancing the phospholipid-dependent PKC activation. PKC activation and subsequent translocation to membrane compartments is a key event in the PKC-dependent signaling cascade, and is preceded by agonist-mediated stimulation of receptor-associated phospholipase C (PLC). PLC, upon stimulation, cleaves inositol-1,4,5-trisphosphate and diacylglycerol from the membrane-bound phosphatydylinositol moiety. In addition, it was demonstrated that RACK may work in concert with the PKC activation process both by directing activated PKC molecules to specific subcellular compartments and stabilizing their active state within these particular compartments.3
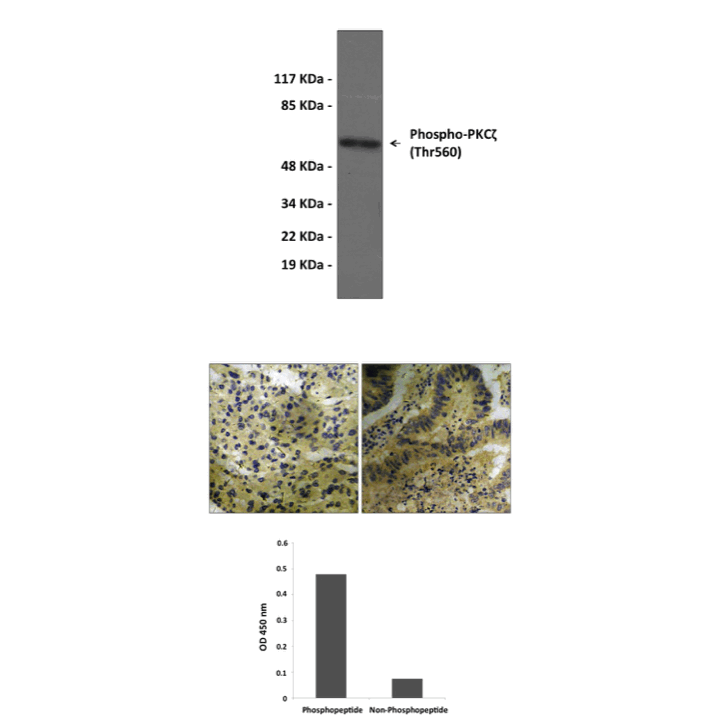
There is numerous evidence that PKC activity is under control by phosphorylation. It was shown that three catalytic domain ‘priming’ phosphorylation sites are important for PKC activity, i.e. the activation loop site, the autophosphorylation site and the hydrophobic, C-terminal site. PDK1 plays a major role in phosphorylation of the activation loop site. When phosphorylated in these three priming sites, PKC has a higher specific activity than when unphosphorylated . Thus, DAG (+Ca2+) will acutely switch these transducers on (c/nPKC isotypes), but the volume control is a function of phosphorylation. Thus, phosphorylation occurs efficiently in vivo when PKC is in an active, i.e. effector-bound, conformation. For PKC, the consequence of phosphorylation is an increase in latent catalytic activity, without bypassing the requirement for allosteric activators.4 Thr410 is in activation loop of PKC zeta was phosphorylated by PDK1 and Thr560 is a autophorylation site. Phosphorylation of both sites potentiate PKC zeta kinase activity. However, it was also reported that PKC-zeta activity is downregulated through proteolysis after phosphorylation at Thr410 and autophosphorylation at Thr560.5
RECENT PUBLICATION
An, K., C. Zhen, Z. Liu, Z. Zhao, H. Liu, X. Zhong, and W. Huang. 2015. Spinal protein kinase Mζ contributes to the maintenance of peripheral inflammation-primed persistent nociceptive sensitization after plantar incision. European Journal of Pain, 19:29-47.
REFERENCES
1. Ohno, S. & Nishizuka, Y.: J. Biochem. 132:509-11, 2002
2. Messerschmidt, A. et al: J. Mol. Biol. 352:918-31, 2005
3. Webb, B.L.J. et al: Br. J. Pharmcol. 130:1433-52, 2000
4. Parekh, D.B. et al: EMBO J. 19:496-503, 2000
5. Sajan, M.P. et al: Am. J. Physiol. Endocrinol. Metab. 298:E179-E192, 2009
Products are for research use only. They are not intended for human, animal, or diagnostic applications.
Cat.No.: | CG1453 |
Antigen: | Range AA545 to 575 |
Isotype: | Rabbit IgG |
Species & predicted species cross- reactivity ( ): | Human, Rat, Mouse |
Applications & Suggested starting dilutions:* | WB 1:500-1:1000 IP n/d IHC 1:50-1:100 ICC n/d FACS n/d ELISA 1:20000 |
Predicted Molecular Weight of protein: | 67 kDa |
Specificity/Sensitivity: | Detects endogenous PKCζ proteins without cross-reactivity with other family members. |
Storage: | Store at -20°C, 4°C for frequent use. Avoid repeated freeze-thaw cycles. |
*Optimal working dilutions must be determined by end user.